The architecture of striated muscle consists of a number of components that work in conjunction to generate a contraction. If you segment a muscle into its most basic component, the sarcomere, you can begin to understand the fundamental processes that take place to ultimately allow us to move limbs, breathe or pump blood through the body. In simplest terms, a sarcomere consists of the myofilament proteins myosin and actin, commonly referred to as the thick and thin filament.
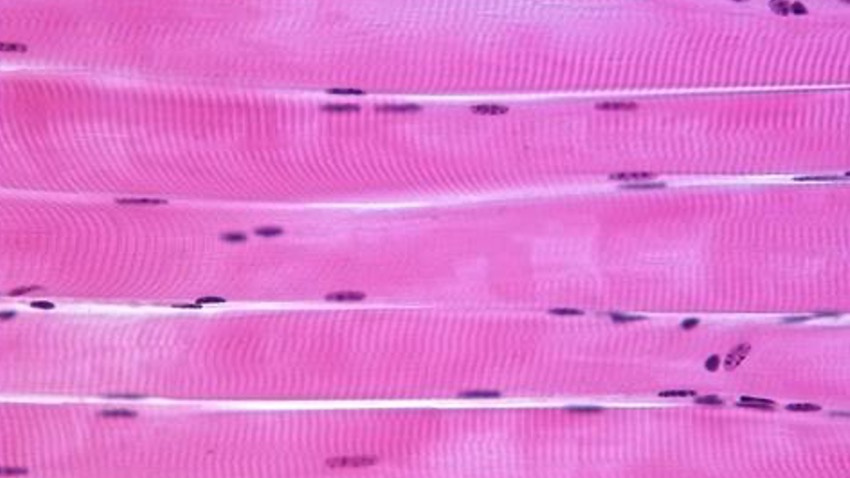
As shown in Figure 1, the sarcomere can be segmented into different regions based on the protein arrangement. As you can see, the I band contains actin only, the A band has overlapped actin and myosin filaments, and the M band is the middle of the H zone where no actin is present. When viewed under a microscope, you will notice a series of dark vertical bands that span an individual cell or fiber. These dark bands are the z-disc region of a sarcomere which represent the borders of each individual sarcomere unit. The distance from z-disc to z-disc is used to measure sarcomere length (SL).At this point you may be thinking, why does the distance between z-discs matter? It is important to understand that when calcium flows into a cell, it leads to activation of the myosin heads along the thick filament which in turn bind to actin, forming a cross bridge. When these cross bridges are formed, the thick and thin filament slide past one another based on the sliding filament theory. The summation of each sarcomere unit shortening is what produces contraction in a cell, fiber and ultimately the muscle. Within the A band, the region of overlap between the thick and thin filaments can vary depending on the sarcomere length. At any one particular SL or fiber length, there’s a defined number of myosin heads available to bind to actin and generate tension. The maximum force generated by a muscle is thus dependent on the starting length of the sarcomere and this region of overlap.
The mechanism behind this is based on a Length-Tension curve which is illustrated in Figure 2. When there’s an overly long SL as shown at point (5), the muscle generates little to no force as myosin and actin do not have any overlap within the A band and therefore there are no myosin heads available to attach to actin to form a cross bridge. As you decrease SL from (5 to 4), a muscle can generate greater and greater force as the overlapping area between myosin and actin gets larger, permitting a greater number of cross bridges to be formed. This rise in force continues until you reach a plateau (3) where the leading edge of the thin filament encounters a bare region of myosin heads and has formed as many cross bridges as possible. Therefore, this region is referred to as the optimal resting length where maximum force is generated. As SL continues to decrease (3 to 2), actin filaments begin to overlap, leading to interference with cross-bridge formation and loss of tension. Further shortening of SL (2 to 1), force generation quickly drops to zero on the grounds that interference of cross-bridge formation reaches its peak and myosin heads block further movement of the two actin edges, thereby inhibiting further shortening.It is therefore very important to take this Length-Tension relationship into account when characterizing muscle mechanics and function in isolated preparations like single cell, fiber or bundle. Setting optimal resting length by utilizing a sarcomere length measurement system ensures your preparation is standardized between each individual tissue so you’re generating maximum tension each and every time. This also provides the ability to produce Length-Tension curves at an SL level to directly compare myofilament properties between experimental groups.